Phage Cro Protein

|
A. Introduction to Phage Cro Protein
B. Crystal Structure of Phage Cro Protein
C. Phage Cro Proteins's Involvement in Bacteriophage Infection
D. Phage Cro as a Therapeutic Agent
|
An Introduction to Phage Cro Protein
Phage Cro Protein is involved in transcription, the process of copying DNA's genetic information into RNA so that it can be made into proteins, and DNA's instructions can be carried out within the cell. The University of Nebraska has a good animation that describes the process of transcription in more detail (although the animation describes transcription in a plant cell, the concept is essentially the same).
Cro is a negative regulator. That is, it works to repress the transcription of other proteins (primarily, lambda repressor) by binding preferentially to the operator site that partially covers lambda repressor's promoter region. The mechanism by which Cro works, alongside lamda repressor, is perhaps the most well known (Degnan et al, 2007). Additionally, other phages utilize essentially the same mechanism. The regions in which Cro and lambda repressor are found are conserved throughout most of the other phages (Degnan et al, 2007).
It has even been suggested that the nonspecific binding by Cro protein can also play an affect within the cell (Bakk and Metzler, 2004). In fact, for relatively high concentrations of Cro, upwards of half of the protein is bound nonspecifically (Bakk and Metzler, 2004). Bakk and Metzler suggest that the nonspecific binding of Cro and lambda repressor is important for maintaing the stability of the genetic switch (Bakk and Metzler, 2004).
Crystal Structure of Phage Cro Protein
Cro's structure consists of 2 beta sheets and 3 alpha helices, of which the helices are arranged in a helix-turn-helix conformation, as shown in the picture above. It is thought that the Phage Cro protein, like other Cro Proteins, are descended from an all-helical ancestor (Dubrava et al, 2008). Cro uses the helix-turn-helix motif to bind to the half site on the DNA, which is very close to the consensus sequence. One of the helices, the recognition helix (a3) is able to insert itself within the major groove of the DNA and interact with the bases.
The Online Macromolecular Museum has an excellent exhibit, which explains Cro's binding process in greater detail.
When Cro binds to its operator site, it binds as a dimer, which gives it extra stability. The dimer forms due to both hydrophobic interactions, as well as interactions between the beta sheets of the protein. Lambda Cro dimerizes in solution, and has a relatively low dissociation constant (LeFevre and Cordes, 2003). Additionally, the dimer is much more themodynamically stable than the monomer--the difference in melting temperatures has been shown to be up to 30 degrees (LeFevre and Cordes, 2003).
The Lambda Cro protein, consisting of both alpha and beta units, evolved from an entirely alpha protein (LeFevre and Cordes, 2003). Previously, one of the amino acids used for dimerization (and Alanine) was actually a much more hydrophobic tryptophan (LeFevre and Cordes, 2003). As discussed by Newlove et al, it is likely that the switch occured by homologous switching as opposed to nonhomologous switching (Newlove et al, 2004)
Phage Cro's Involvement in Bacteriophage Infection
When bacteriophages first inject their DNA into bacteria cells, the first genes to be transcribed are for Cro and N protein. As a negative regulator, Cro works to suppress the production of lambda repressor, which (if turned on) would repress Cro, and cause the newly created prophage to enter lytic mode. If Cro stays on, the cell enters the lytic mode--since lambda repressor is turned off, Q protein (activated by N protein) acts as an antiterminator and stimulates the transcription of the proteins needed for lysis (to make the head and tail of the protein coat, as well as for the actual lysing of the bacteria cell).
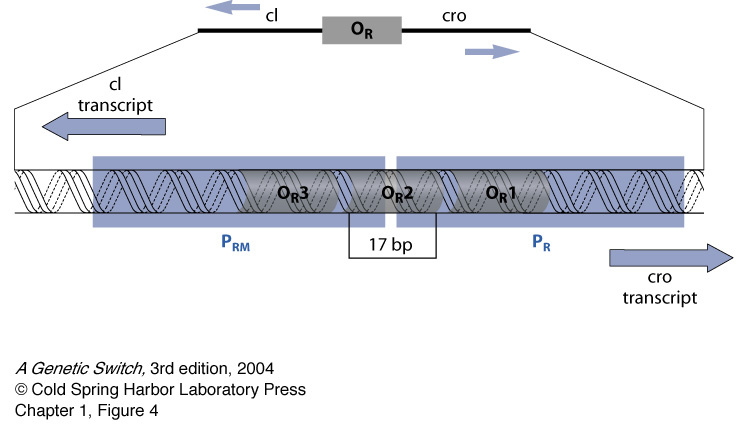
As stated before, Cro and Lambda Repressor work in opposition to each other-Cro suppresses the transcription of Lambda Repressor, while Lambda Repressor both represses Cro and activates transcription of itself. When only Lambda Repressor is present in the system, it will bind to Operator 1 (OR1), which partially covers Cro's promoter region (see below, to see how). Because it displays cooperative binding, another Lambda Repressor is able to quickly bind to Operator 2 (OR2), even though it has far less affinity for it as compared to OR1. A Lambda Repressor on OR2 is able to interact with the RNA Polymerase when it sits down on Lambda Repressor's promoter region, activating transcription and keeping the cell in lysogenic mode.

However, it is possible for there to be too little Lambda Repressor in the system for it to accurately bind to OR1 and OR2-generally due to the transformation of RecA into a protease by UV radiation, causing it to degrade Lambda Repressor (www.microbiologybytes.com). In this case, Cro is able to bind to Operator 3 (OR3), which partially covers Lambda Repressor's promoter region, and shut it off. Because Cro's promoter region is closer to the consensus sequence for the sigma factor of RNA Polymerase, the Polymerase is able to quickly transcribe large amounts of Cro, effectively switching the cell into lytic mode (www.microbiologybytes.com).
Schubert et al performed an experiment in which they examined the true purpose of Cro's role in the genetic switch by mutating OR3 (Schubert et al, 2007). Although the mutation did not effect the phage's development after infection, it kept the phage from switching from lysogenic to lytic mode (Schubert et al, 2007). Even upon the addition of UV light, leading to the degradation of Lambda Repressor, the cell did not switch from lysogeny to lysis-more Lambda Repressor was synthesized instead (Schubert et al, 2007).
Another, more recent, study by Cao et al showed that as Cro levels increase dramatically (and lambda repressor levels drop), the prophage undergoes a rapid switch from lysogenic to lytic cycles, although mutants don't undergo the same dramatic switch (2008).
An animation which describes this switch in some detail can be found here.
Phage Cro Protein as a Therapeutic Agent
1. As discovered in an experiment performed by Facchini and Lingwood, Cro protein could potentially aid in the incorporation of foreign DNA into cells (Facchini and Lingwood, 2001). Their experiments involved using another protein, a verotoxin subunit, which was bound to the Cro protein, which therefore has the ability to bind to not only to Cro's operator, but also glycolipids on the surface of the cell (Facchini and Lingwood, 2001). The DNA has been shown to be incorporated successfully into several different organelles within the cell (Facchini and Lingwood, 2001). This discovery could eventually be used for gene therapy.
2. Parera et al have discovered a method of detecting SARS (Severe Acute Respiratory Syndrome) that utilizes the genetic switch involving Cro and Lambda Repressor (Parera et al, 2004). The agent involved in SARS, a coronavirus, utilizes site specific degradation of proteins (Parera et al, 2004). The target sequence for the virus was inserted into the lambda repressor (Parera et al, 2004). After infection, the E. coli cells into which the virus was inserted showed a dramatic increase in the number of new viruses that were created as compared to E. coli cells infected without the insert (Parera et al, 2004). This is a new, novel method of detecting SARS.
3. Like SARS, Hepititis C's method of attack utilizes a protease-NS3 serine protease (Martinez and Clotet, 2003). The target sequence for the protease was inserted into lambdra repressor (Martinez and Clotet, 2003. After infection, the cells into which the phage with the sequence was placed showed an effienciency of almost 10000 times that of the cells without the insert (Martinez and Clotet, 2003). Like the study of SARS, this is a new way of detecting Hepititis C.
Comments (1)
Christopher Korey said
at 3:28 pm on Apr 6, 2009
Really nice job here. Now transform the outline in to a concise text with links out to more detailed information. Separate each subsection by inserting a horizontal line. I would probably get rid of the second image unless there is an important reason to have it there--maybe link out to the page it came from if you want to include it that way. I like the biotech/therapeutic things you found, very cool. In the footnotes (3-5) put a regular citation in there and then make it a link with the url you have pasted in. This will take them directly to the abstract and it looks cleaner.
You don't have permission to comment on this page.